ο Προκλητική εφαρμογή-Η θεωρία επιστρέφει στο Koshland και αντιστοιχεί σε μια επέκταση της αρχής της κλειδαριάς-κλειδώματος, η οποία βασίζεται στην ακρίβεια της προσαρμογής των ανατομικών δομών. Η επαγόμενη εφαρμογή αναφέρεται σε ένζυμα όπως η κινάση που αλλάζουν τη διαμόρφωσή τους για να σχηματίσουν ένα σύμπλοκο ενζύμου-προσδέματος. Στην περίπτωση ελαττωμάτων ενζύμου, η αρχή της επαγόμενης προσαρμογής μπορεί να επηρεαστεί από διαταραχές.
Τι είναι το Induced Fit;

Πολλές διαδικασίες στο σώμα λειτουργούν με την αρχή του κλειδώματος κλειδιού ή του χεριού-γάντι. Αυτό ισχύει, για παράδειγμα, για αρθρωτές συνδέσεις. Η κεφαλή της άρθρωσης εμπλέκεται στην υποδοχή της άρθρωσης σαν ένα κλειδί σε μια κλειδαριά ή ένα χέρι σε ένα γάντι. Η πόρτα ανοίγει μόνο όταν το κλειδί βρίσκεται ακριβώς στην κλειδαριά. Στο ίδιο πλαίσιο, ορισμένες λειτουργίες του σώματος ανοίγουν μόνο όταν οι δομές συναντώνται ακριβώς.
Το Induced-fit είναι μια ειδική μορφή της αρχής κλειδώματος-κλειδώματος. Αυτή είναι μια θεωρία για τον σχηματισμό συμπλοκών πρωτεϊνών-συνδετών, για παράδειγμα ενός συμπλόκου ενζύμου-υποστρώματος στο πλαίσιο αντιδράσεων που καταλύονται από ένζυμα.
Ο Daniel E. Koshland θεωρείται ο πρώτος που περιέγραψε τη θεωρία και το διατύπωσε για πρώτη φορά το 1958. Σε αντίθεση με την αρχή κλειδαριάς-κλειδώματος, η θεωρία της επαγόμενης προσαρμογής δεν προϋποθέτει δύο στατικές δομές. Ειδικά στην περίπτωση συμπλοκών πρωτεΐνης-συνδετήρα, μια διαμορφωτική αλλαγή στην εμπλεκόμενη πρωτεΐνη θα πρέπει να επιτρέψει το σχηματισμό του συμπλόκου. Συνδέτης και πρωτεΐνη, ή μάλλον ένζυμο, ο Koshland θεωρούσε δυναμική και μίλησε για μια αλληλεπίδραση που μετακινεί και τους δύο εταίρους σε μια αλλαγή στη διαμόρφωση για χάρη του σύνθετου σχηματισμού.
Λειτουργία & εργασία
Υπάρχει δεσμευτική ειδικότητα μεταξύ ενζύμων και υποστρωμάτων. Αυτή η δεσμευτική ειδικότητα υπονοεί την αρχή κλειδώματος κλειδιού. Κάθε ένζυμο έχει ένα ενεργό κέντρο. Για σύνθετο σχηματισμό με πρόσδεμα, αυτό το κέντρο έχει ακριβώς σχήμα έτσι ώστε να ταιριάζει σχεδόν τέλεια με το χωρικό σχήμα του επιδιωκόμενου υποστρώματος.
Στην περίπτωση πολλών ενζύμων, ωστόσο, το ενεργό κέντρο σε κάθε περίπτωση είναι σε μορφή που δεν είναι πολύ ακριβής εφόσον δεν δεσμεύεται σε ένα υπόστρωμα. Αυτή η παρατήρηση φαίνεται να έρχεται σε αντίθεση με την αρχή κλειδώματος και κλειδιού, επειδή τα ένζυμα και τα προσδέματά τους αρχικά φαίνεται να προσαρμόζουν το σχήμα τους.
Μόλις το ένζυμο προσκολληθεί σε ένα πρόσδεμα, δημιουργούνται διαμοριακές αλληλεπιδράσεις. Αυτές οι αλληλεπιδράσεις σε διαμοριακό επίπεδο οδηγούν σε αλλαγή στη διαμόρφωση του ενζύμου. Η διαμόρφωση εννοείται ότι σημαίνει τις διάφορες πιθανές διευθετήσεις μεμονωμένων ατόμων σε ένα μόριο που προκύπτει από μια απλή περιστροφή γύρω από έναν άξονα. Η αλλαγή στη διαμόρφωση των ενζύμων αντιστοιχεί σε μια αλλαγή στη χωρική διάταξη των μορίων τους και επιτρέπει μόνο τον σχηματισμό ενός συμπλόκου ενζύμου-υποστρώματος.
Η εξακινάση ως ένζυμα καταλύει, για παράδειγμα, το πρώτο βήμα της γλυκόλυσης. Μόλις αυτά τα ένζυμα έρθουν σε επαφή με τη γλυκόζη του υποστρώματος, μπορεί να παρατηρηθεί μια επαγόμενη προσαρμογή με την έννοια του σχηματισμού «επαγόμενης προσαρμογής». Το ένζυμο εξακινάση φωσφορυλιώνει τη γλυκόζη υποκαταστάτη του καταναλώνοντας ΑΤΡ για να σχηματίσει 6-φωσφορική γλυκόζη.
Η δομή του νερού μοιάζει με εκείνη στην αλκοολική ομάδα του ατόμου C6, την οποία το ένζυμο φωσφορυλιώνει κατά τη διάρκεια της αντίδρασης. Λόγω του μικρού μεγέθους, τα μόρια νερού θα μπορούσαν να προσκολληθούν στο ενεργό κέντρο του ενζύμου, έτσι ώστε να δημιουργηθεί υδρόλυση του ΑΤΡ. Ωστόσο, η επαγόμενη εφαρμογή επιτρέπει στην εξωκινάση να καταλύει τη μετατροπή γλυκόζης με υψηλή ειδικότητα, έτσι ώστε η υδρόλυση ΑΤΡ να πρέπει να πραγματοποιηθεί σε μικρό βαθμό. Με τον μηχανισμό επαγόμενης προσαρμογής, αυξάνεται η ειδικότητα του υποστρώματος.
Η αρχή του ανθρώπινου οργανισμού μπορεί να τηρηθεί ιδιαίτερα στην περίπτωση των κινασών. Η επαγόμενη προσαρμογή δεν ισχύει για κάθε σύμπλοκο προσδέματος-υποδοχέα, καθώς η διαμορφωτική αλλαγή και των δύο εταίρων είναι σε πολλές περιπτώσεις φυσικά όρια.
Ασθένειες και παθήσεις
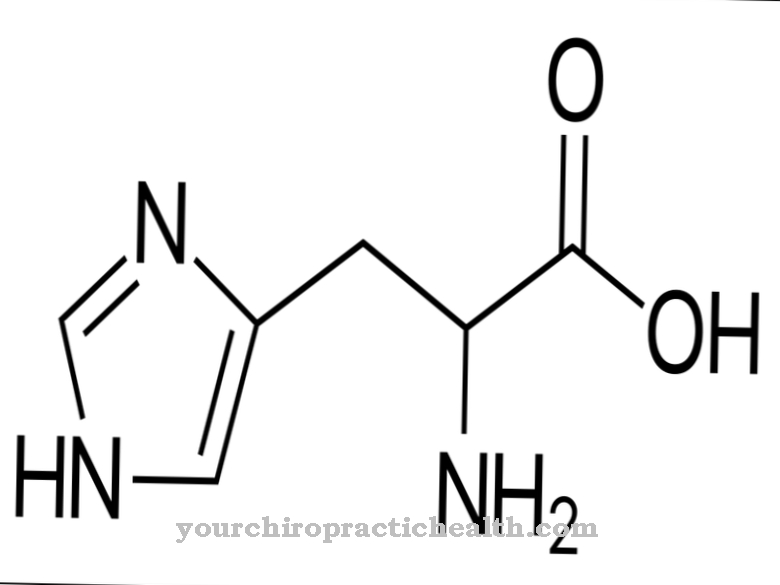
Η αρχή της επαγόμενης προσαρμογής διαταράσσεται στην περίπτωση διαφορετικών ενζυματικών ελαττωμάτων. Στη φαινυλκετονουρία, για παράδειγμα, τα ένζυμα περιορίζονται στη δραστηριότητά τους ή αποτυγχάνουν εντελώς. Συνήθως αυτό οφείλεται σε γενετικό ελάττωμα. Στη φαινυλκετονουρία, το ένζυμο υδροξυλάση φαινυλαλανίνης είναι ελαττωματικό. Η φαινυλαλανίνη δεν μετατρέπεται πλέον σε τυροσίνη και συσσωρεύεται ανάλογα. Εμφανίζονται νευροτοξικές ουσίες, έτσι ώστε εκτός από τις ψυχικές αναπηρίες, ο ασθενής έχει την τάση να κράμπει. Οι ενζυματικές ανωμαλίες είναι συνήθως γενετικές και προκαλούνται από εσφαλμένα κωδικοποιημένη αλληλουχία αμινοξέων στο DNA.
Οι μεταβολικές ασθένειες που προκαλούνται από ενζυματικές ατέλειες και μια τέτοια διαταραγμένη αρχή της επαγόμενης προσαρμογής είναι γνωστές ως ενζυμοπάθειες. Τα ελαττώματα πυροσταφυλικής κινάσης υπάρχουν, για παράδειγμα, σε ένα εσφαλμένα κωδικοποιητικό γονίδιο PKLR. Αυτό το γονίδιο βρίσκεται στον γονιδιακό τόπο 1q22 του χρωμοσώματος 1. Είναι γνωστές διάφορες μεταλλάξεις του αλληλόμορφου PKLR της πυροσταφυλικής κινάσης, οι οποίες εμφανίζονται ως ελαττώματα στη μορφή R.
Η ασθένεια Hers αναφέρεται και πάλι ως γλυκογένεση τύπου VI και ανήκει στην ομάδα ασθενειών αποθήκευσης γλυκογόνου. Είναι μια αυτοσωματική υπολειπόμενη ή συνδεδεμένη με Χ μεταβολική διαταραχή λόγω ενζυματικών ελαττωμάτων. Πιο συγκεκριμένα, η αιτία έγκειται σε διαφορετικά ενζυματικά ελαττώματα στο σύστημα φωσφορυλάσης κινάσης εντός του ήπατος και των μυών. Γνωστό σε αυτό το πλαίσιο είναι, για παράδειγμα, το ελάττωμα φωσφορυλάσης β-κινάσης συνδεδεμένης Χ στο ήπαρ, το ελάττωμα φωσφορυλάσης ήπατος στην αυτοσωμική υπολειπόμενη κληρονομικότητα και η συνδυασμένη αποτυχία της φωσφορυλάσης β-κινάσης εντός του ήπατος και των μυών.
Σε σχέση με τη φωσφορυλάση του ήπατος, οι αιτιολογικές μεταλλάξεις εντοπίστηκαν στο γονίδιο PYGL και επομένως βρίσκονται στο χρωμόσωμα 14q21 έως q22. Η συνδυασμένη ανεπάρκεια φωσφορυλάσης των ηπατικών μυών έχει συνδεθεί με μεταλλάξεις στο γονίδιο PHKB στον τόπο 16q12-q13. Αιτιώδεις μεταλλάξεις στο γονίδιο PHKA2 στον τόπο Xp22.2-p22.1 εντοπίστηκαν για το X-συνδεδεμένο ελάττωμα στην κινάση φωσφορυλάσης του ήπατος. Άλλες γλυκογονόζες μπορούν επίσης να ακυρώσουν το επαγόμενο αποτέλεσμα προσαρμογής της αντίστοιχης κινάσης ή να το κάνουν πιο δύσκολο.














.jpg)

.jpg)